師部

師部(篩部、しぶ、英: phloem)とは、維管束植物(シダ植物や種子植物)の維管束を構成する2つの複合組織のうちの1つであり(もう1つは木部)、師管(篩管、しかん)や師細胞組織、師部繊維組織、師部柔組織からなる。頂端分裂組織に由来する一次師部(図1a)と、維管束形成層に由来する二次師部(靭皮; 図1b)がある。師部の主な機能は光合成産物など有機物の輸送であり(図1c)、また植物体の機械的支持や物質貯蔵などにも寄与している。

.jpg)

「師部」の名は、師管の隔壁が篩(ふるい)のような形であることに由来するため、本来は「篩部」であり、また関連する「師管、師板、師細胞」なども本来は全て「篩管、篩板、篩細胞」などである。しかし「篩」が常用漢字ではないため(2020年現在)、教科書などでは「師」で代用される(ただし「師」には「ふるい」の意味はない)[1]。木(木本植物)において師部は樹皮の内層に存在しており、師部を意味する英語の phloem は樹皮を意味するギリシア語の φλοιός (phloios) に由来する[2]。
構成
[編集]師部は、師要素(師管要素や師細胞)、師部繊維、師部柔細胞からなる[3][4][5]。
師要素
[編集]師要素(篩要素、しようそ、sieve element)は管状の細胞であり、光合成産物などの有機物の通道に機能している[3][5](→#師部輸送(師部転流)を参照)。師要素は生細胞ではあるが、核や液胞などの細胞小器官がしばしば退化しており[3]、物質が通道しやすくなっている[6]。またしばしば特殊な色素体をもつ[7]。細胞壁はやや厚いが、二次細胞壁を欠く[3][5][8]。細胞間の物質輸送は、原形質連絡が拡大した師孔(篩孔、sieve pore)を通して行われる[3][5][9]。師孔において、原形質連絡の周囲はカロースが沈着(肉状体ともよばれる)している[7][9]。師要素の細胞表面には、多数の師孔が集まって師域(篩域、しいき、sieve area)を形成している[3][5][10]。師要素が機能するのはふつう数ヶ月程度であり(長年機能している例もある)、一定期間後または損傷した師要素ではカロースがさらに沈着して師孔は閉塞される[3][8][9]。師要素として、被子植物は師管要素を、それ以外の維管束植物(シダ植物や裸子植物)は師細胞をもつ[11][5]。


師管要素(篩管要素、しかんようそ、sieve tube element, sieve tube member; 師管細胞、sieve tube cell)は、上下端で縦につながって師管(篩管、しかん、sieve tube)を形成している[3][5][11](図2a)。師管要素どうしの上下の隔壁は師板(篩板、しばん、sieve plate)とよばれ、大型の師孔(ときに直径 10 µm 以上)が集合した「ふるい(篩)」のような師域が形成されている[3][5][8][10][11](図2b)。師板における師域の配置様式には多様性があり、細長い師域が階段状に配置しているもの(例: シュウカイドウ)や不定形の師域が集まっているもの(例: トウ属)は複合師板 (compound sieve plate) とよばれ、1個の大きな師域をもつもの(例: カボチャ; 図2b)は単師板 (simple sieve plate) とよばれる[11][12][13]。師管における物質輸送は、主に師板を通して行われるが、ふつう側面にも師域が存在する(側師域)[12]。ただしこのような師域の師孔は小さい[8][11]。師板は側壁に対して斜めのものからほぼ直角のものまである[8][12]。師管要素には、同一の母細胞から不等分裂によって形成された小型の細胞である伴細胞(はんさいぼう、companion cell)が付随している[3][5][7][11][14]。伴細胞は多数の原形質連絡(ときに原形質連絡が分枝)によって師管要素とつながっており、糖などは伴細胞を介して師管要素へ運ばれる[6][8]。また伴細胞はタンパク質などを送ることで師管要素の代謝を制御していると考えられている[6][14]。師管要素・師管は、基本的に被子植物に特徴的な構造であるが、被子植物の中でアウストロバイレヤ科は師管を欠く[15]。
師細胞(篩細胞、しさいぼう、sieve cell)は一般的に師管要素より細長く紡錘形であり、側面で互いに接して師細胞組織(篩細胞組織、sieve cell tissue)を形成している[16]。師板をもたず、細胞側面に散在する師域を通して物質輸送が行われる[7]。師管にくらべて通道効率は低いと考えられている[16]。師細胞には伴細胞は付随していないが、裸子植物にはタンパク細胞 (albuminous cell) とよばれる細胞が師細胞に付随しており(師細胞と姉妹の関係にはない)、伴細胞と同様に師細胞の機能を補助していると考えられている[3][17]。師細胞は、被子植物以外の維管束植物に存在する。
師管要素と師細胞の関係は、木部における道管要素と仮道管の関係に似ているため、師管を道管状師管 (vessel form sieve tube)、師細胞組織を仮道管状師管 (tracheid form sieve tube) とよんでいたこともある[11][16]。
師部繊維
[編集].jpg)
師部繊維(篩部繊維、phloem fiber)は発達した細胞壁をもつ細長い細胞であり、ふつう原形質を欠く死細胞である[5]。師部繊維は、集合して師部繊維組織(篩部繊維組織、phloem fiber tissue)を形成している[5](図3)。師部繊維は、木部繊維などと共に、植物体の機械的支持に寄与している。師部繊維は木部繊維にくらべて一般的に長く、またふつう壁孔は不明瞭である[5]。長さはアサ(アサ科)で 1 cm、アマ(アマ科)で 2–4 cm、カラムシ(イラクサ科)では 22 cm に達する。これらの繊維は布や縄の原料として利用される(下記参照)。師部繊維は、ふつう師要素の外側に形成される[18](図3)。また維管束周辺に厚壁細胞が発達する種では、師部繊維は発達しない[19](例: カボチャ)。被子植物以外では、ふつう師部繊維は見られない[5]。
師部柔細胞
[編集].jpg)
師部に存在する柔細胞は師部柔細胞(篩部柔細胞、phloem parenchymatous cell)をよばれ、 集合して師部柔組織(篩部柔組織、phloem parenchyma)を形成している[5]。有機養分の貯蔵などに働いている[5]。タンニンや樹脂を貯蔵するものや分泌組織をもつこともある[5][8]。一次師部(下記)では、師部柔細胞は師要素に付随して存在する[4]。二次師部(下記)の場合には、二次木部の柔組織と同様、長軸にならんだ細胞からなる軸方向柔組織 (axial parenchyma) と、放射方向に配列した細胞からなる放射柔組織 (ray parenchyma) がある[5](図4)。
一次師部と二次師部
[編集]師部は、形成される場所に応じて一次師部と二次師部に分けられる[5][19][20]。
一次師部
[編集]頂端分裂組織(シュート頂分裂組織と根端分裂組織)に由来する前形成層 (procambium) から形成される師部は、一次師部 (primary phloem) とよばれる[19][21]。一次師部は、ふつう一次木部の間(図5a)や外側(図5b)に位置するが、師部が木部を取り囲んでいる例もある[4](図5c)。全ての維管束植物は一次師部をもつが、二次成長を行う木本植物では、二次師部の発達と共に一次師部は破壊されていく[19]。
.jpg)
.jpg)

一次師部のうち、最初に分化する部分を原生師部 (protophloem)、その後に分化する部分を後生師部 (metaphloem) とよぶ[5][19][21]。原生師部・後生師部の形態的な差異は、原生木部・後生木部ほど明瞭ではないが、一般的に後生師部のほうが太くて長い[19]。また原生師部の師管には伴細胞が見られないことが多い[19]。
二次師部
[編集]茎や根において、一次師部と一次木部の間に生じた分裂組織である維管束形成層(単に形成層とよばれることも多い)から形成された師部は、二次師部(次生師部、secondary phloem)とよばれる[5][20]。二次師部は、維管束形成層の外側に付加されていく[20][22](図6)。維管束形成層による二次成長を行う植物(いわゆる木本植物)は二次師部をもち、このような植物では二次師部は樹皮の内層に位置する。生物学における靭皮(じんぴ、bast)は、二次師部のことを意味する[5]。二次師部では、一次師部にくらべて構成要素が放射方向に整然と並んでいる傾向があり、また師要素が短く密度が高い[20]。

.jpg)
.jpg)
二次師部のそれぞれの師要素が機能するのは短期間であり、内側から付加される新たな組織に押し出され、崩壊していく。そのため、二次木部の厚さが年々増加していくのに対して、二次師部の厚さはある程度以上は変化しない[5][8]。
二次木部と同様、二次師部にも軸方向の要素(師要素、師部繊維、軸方向柔組織)と放射方向の要素(放射柔組織)が存在する[23]。これらの始原細胞は維管束形成層に存在し、二次木部と共有している。そのため、二次師部の放射組織は、維管束形成層を挟んで二次木部の放射組織と連続している[8][23](図6c)。また二次成長している茎や根は円周を増大させていくため、師部放射組織はそれを埋めるよう細胞分裂によって外側に向かって幅広くなっていることがある(図6c)。このような放射組織は、拡張放射組織 (dilated ray) とよばれる[7]。
師部輸送(師部転流)
[編集]光合成によってつくられた糖は、師部の師要素からなる師管や師細胞組織を通して植物体全体に輸送される(図7)。この輸送は、師部輸送(篩部輸送、phloem transport)または師部転流(篩部転流、phloem translocation)とよばれる[24][25]。師部輸送によって運ばれる水溶液には大量の糖(主にスクロース、ときにオリゴ糖または糖アルコール)が含まれ、またアミノ酸や無機養分も含まれている[24]。さらに植物ホルモンや伝令RNA (mRNA) も輸送され、植物体全体の情報交換の通路ともなっている[24][26]。このような師部輸送によって運ばれる液は、師管液(篩管液、phloem sap)とよばれる[26]。師部輸送では、師管断面積 1 cm2 あたり1時間に数グラムの物質が輸送され、また輸送速度は時速 1 m に達することがある[24]。
木部による水や無機養分の輸送(木部輸送)が根から気孔がある部分へ向けての一方通行の輸送であるのに対して、師部輸送の方向は時期や植物の状態によって変動する[26]。このような師部輸送において、師管液を供給する側をソース (source)、師管液を受け取る側をシンク (sink) という[26]。同一の器官が時期によってソースとなったりシンクとなったりすることがあり、例えば貯蔵器官は光合成が活発に行われている時期にはシンクとなり、早春など活発な活動を開始する前にはソースとなる。シンクとソースの間の師部輸送の仕組みは、基本的にシンクよりソースの方が浸透圧が高いことによって生じる水の流れであると考えられている。この仮説は圧流説 (pressure flow theory) とよばれ、エルンスト・ミュンヒ (1930) によって提唱された[24][26]。ただし被子植物以外の師部輸送(師管ではなく師細胞組織が用いられる)が、圧流説で説明できるか否かは明らかではない[26]。
ソースから師部に物質が移動することは積み込み(師部積み込み、ローディング、phloem loading)とよばれ、師部からシンクに物質が移動することは積み降ろし(師部積み下ろし、アンローディング、phloem unloading)とよばれる[24][25][26]。いずれの場合も、シンプラスト経路(原形質連絡を通した経路)とアポプラスト経路(細胞壁など細胞膜外を通した経路)が存在する[24][26]。例えば原形質連絡を通じて運ばれたスクロースが中継細胞 (intermediate cell) においてオリゴ糖に変換され、オリゴ糖は大きいため葉肉細胞へは戻れず、師管方向への輸送が促進される植物もある(ポリマートラッピング polymer trapping)[24]。また積み込みの際には、糖の濃度が低いところから高いところへ輸送されることがあり、このような場合は能動輸送も用いられる[26]。例えばスクロースが能動輸送によって輸送細胞 (transfer cell) に取り込まれ、これが師管へ輸送されるものもある[24]。積み降ろしの場合は、シンク側で糖が消費またはデンプンなどの不溶性物質に変換されているため、ふつうシンク側の方が糖濃度が低くなっている[26]。
師部を通した電気的なシグナルの伝達も知られている。植物体のある部分で生じた電気刺激が師部を通して伝達され、遺伝子発現や呼吸、光合成、植物ホルモンの量、師部での積み降ろし(上記)などに変化を引き起こすことが報告されている[26]。
利用
[編集]繊維など
[編集]

師部繊維は、その外側に形成される皮層繊維と共に靭皮繊維 (bast fiber) とよばれ、天然繊維としてさまざまな用途に用いられている[18]。アマ(リネン、アマ科)、アサ(麻、アサ科)、カラムシ(苧麻、イラクサ科)、ツナソ(ジュート、アオイ科)などの靭皮繊維は布や縄の原料に(図8a)、コウゾ(クワ科)やミツマタ、ガンピ(ジンチョウゲ科)の靭皮繊維は和紙や紙幣の原料に用いられる[27][28](図8b)。
パラゴムノキ(トウダイグサ科)の二次師部に存在する乳管から分泌される乳液(ラテックス)にはさまざまなポリテルペンが含まれ、天然ゴムとして利用される[8]。
飲用・食用・薬用
[編集]

師管液には糖(主にスクロース)などが含まれ、砂糖生産やアルコール飲料の原料に利用されることがある。このような目的では、サトウヤシ、ココヤシ、ナツメヤシ、コウリバヤシ、オウギヤシ、ニッパヤシなどヤシ科の植物がしばしば用いられる[29][30](図9a)。コウリバヤシからは、1日に45リットルもの師管液が採取できる[31]。またメキシコでは、リュウゼツラン属(キジカクシ科)の師管液が甘味料やアルコール飲料(プルケ)の原料として古くから利用されている[32]。
師部には貯蔵物質(糖など)を含む柔細胞を含まれており、フィンランドなど北欧では、飢饉時にマツの靭皮(内樹皮)が救荒食物として利用されていた[33](図9b)。また秋田県の松皮餅もマツの靭皮を利用しており、救荒食物としてつくられたとも伝えられている[34]。ニホンジカなど動物も、ときに木本植物の靭皮を食物として利用しており、これが林業に大きな被害をもたらすこともある[35]。
ケイヒ(クスノキ科)、ホオノキ(モクレン科)、アカメガシワ(トウダイグサ科)、トチュウ(トチュウ科)、キハダ(ミカン科)などでは、靭皮が生薬として用いられている[36]。またキハダの靭皮は、染料としても用いられる[37]。
環状剥皮
[編集]

木本植物において師部は樹皮内層に存在するため、樹皮を環状に剥離することで師部を断ち切ることができ、この手法は環状剥皮(環状はく皮、環状除皮、girdling, ring barking)とよばれる[38](図10a)。環状剥皮は林業、農業、園芸などさまざまな分野で利用される。
例えば樹幹で環状剥皮を行うことによって、その植物を枯死させることができる(巻き枯らし、巻枯らし)。ビーバーはしばしば環状剥皮を行い、移入されたアルゼンチンにおいて樹木を大規模に枯死させたことが報告されている[39]。林業においては、間伐作業の省力化・安全化のために環状剥皮が利用されている[40][41]。ただしこの作業は林内に多数の枯死木を生じさせることになり、幹折れ等による危険や枯死木を利用する害虫増加を招くことが懸念されている。また外来樹種の除去にも同様の手法が利用される[42]。
また枝などに環状剥皮を行うと、その部分より上側に糖など同化産物が留まる。さまざまな樹種において、環状剥皮が樹勢抑制、着花促進、生理落果の抑制、果実品質の向上などに効果的であることが報告されており、ブドウ、カキ、柑橘類などでは実用化されている[43][44][45][46]。また環状剥皮された部分の上で不定根形成が促進されることがあるため、林業、農業、園芸においてこれを使って栄養繁殖させることもある(取り木とよばれる)[47][48](図10b)。
ギャラリー
[編集]-
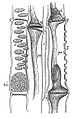
-
 カボチャ属の師部の横断面模式図: 師板 (si) が見える。
カボチャ属の師部の横断面模式図: 師板 (si) が見える。 -
 カボチャ属の師部の縦断面であり、師板が見える。
カボチャ属の師部の縦断面であり、師板が見える。 -
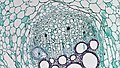 カボチャ属の師部の横断面であり、下部が木部。
カボチャ属の師部の横断面であり、下部が木部。 -
 カボチャ属の師部の横断面: 一部に師板が見える。
カボチャ属の師部の横断面: 一部に師板が見える。 -

-

-

-

-

-

-



.jpg)
.jpg)
.jpg)
.jpg)
.jpg)
.jpg)
.jpg)
.jpg)
.jpg)
.jpg)
脚注
[編集]出典
[編集]- ^ 原 1994, p. 93, 「篩と師」.
- ^ "Phloem". Merriam-Webster.com Dictionary. Merriam-Webster. 2020年4月20日閲覧。
- ^ a b c d e f g h i j k 原 1994, pp. 85–88, 「篩部」.
- ^ a b c 巌佐 et al. 2013, p. 605, 「師部」.
- ^ a b c d e f g h i j k l m n o p q r s t u v 清水建美「師部」『図説 植物用語事典』八坂書房、2001年、185–187頁。ISBN 4-89694-479-8。
- ^ a b c キャンベル 2018, pp. 869–897, 「35 維管束植物の構造、生長、発生」.
- ^ a b c d e ルダル 1997, pp. 28–31, 複合組織.
- ^ a b c d e f g h i j Bowes, Bryan G.; Mauseth, James D. (2008). “Phloem”. Plant Structure: A Colour Guide (2nd Edition ed.). Jones & Bartlett Learning. pp. 67–68. ISBN 978-0763763862
- ^ a b c 巌佐 et al. 2013, pp. 571–572, 「師孔」.
- ^ a b 巌佐 et al. 2013, p. 556, 「師域」.
- ^ a b c d e f g 巌佐 et al. 2013, p. 560, 「師管」.
- ^ a b c アーネスト・M・ギフォード、エイドリアンス・S・フォスター「維管束組織系」『維管束植物の形態と進化』長谷部光泰、鈴木武、植田邦彦 監訳、文一総合出版、2002年、517頁。ISBN 4-8299-2160-9。
- ^ Beck, Charles B. (2005). “Cells of the phloem”. An Introduction to Plant Structure and Development. Cambridge University Press. pp. 33–35. ISBN 978-0521837408
- ^ a b 巌佐 et al. 2013, p. 1119, 「伴細胞」.
- ^ 田村道夫『植物の系統』文一総合出版、1999年、143頁。ISBN 4-8299-2126-9。
- ^ a b c 巌佐 et al. 2013, p. 575, 「師細胞」.
- ^ 柴岡弘郎 (2006年1月23日). “師管について”. みんなのひろば. 日本植物生理学会. 2024年4月29日閲覧。
- ^ a b 巌佐 et al. 2013, p. 713, 「靭皮繊維」.
- ^ a b c d e f g 巌佐 et al. 2013, p. 72, 「一次師部」.
- ^ a b c d 巌佐 et al. 2013, p. 1035, 「二次師部」.
- ^ a b 原 1994, p. 119.
- ^ ルダル 1997, pp. 41–52, 「茎の肥大成長」.
- ^ a b 原 1994, pp. 134–139, 「形成層と二次組織」.
- ^ a b c d e f g h i 巌佐 et al. 2013, p. 606, 「師部輸送」.
- ^ a b L. テイツ、E. ザイガー、I. M. モーラー ほか 編「篩部転流」『テイツ/ザイガー 植物生理学・発生学』(原著第6版)講談社、2017年、288–311頁。ISBN 978-4-06-153896-2。
- ^ a b c d e f g h i j k キャンベル 2018, pp. 899–920, 「36 維管束植物の栄養吸収と輸送」.
- ^ 川鍋祐夫「繊維作物の種類および利用と植物分類上の区分との関係」『熱帯農業』第24巻第2号、1980年、45–53頁、doi:10.11248/jsta1957.24.45。
- ^ 森本正和「非木材繊維利用の現状と将来」『紙パ技協誌』第51巻、1997年、895–914頁、doi:10.2524/jtappij.51.895。
- ^ 佐藤孝「サトウヤシとヤシ砂糖,ヤシ酒」『熱帯農業』第3巻第1号、1959年、6–10頁、doi:10.11248/jsta1957.3.6。
- ^ 荻田信二郎、山口夕「ヤシ科植物のバイオテクノロジー: スリランカとインドネシアの事例」『SAGO PALM』第11巻、2003年、21–25頁、doi:10.57418/sagopalm.11.1_21、NAID 10013952316、 オリジナルの2020年9月19日時点におけるアーカイブ。
- ^ Van Die, J.; Tammes, P. M. L. (1975). “Phloem Exudation from Monocotyledonous Axes”. In Zimmermann, M. H.; Milburn, J. A.. Transport in Plants I. Phloem Transport. Encyclopedia of Plant Physiology New Series, Volume 1. Springer, Berlin. pp. 196–222. ISBN 3-540-07314-0
- ^ 小倉哲也、小嶋良種「アガベから造られるテキーラ、イヌリンなどの製品およびイヌリンの研究経過報告」『FFIジャーナル』第212巻第10号、2007年、872–884頁、NAID 40015693070。
- ^ Lappalainen, M. (2014). “Death and disease during the great Finnish famine 1695-1697”. Scandinavian Journal of History 39 (4): 425–447. doi:10.1080/03468755.2014.937740.
- ^ 三森一司、佐藤奈緒美「松皮に含まれる食物繊維の加熱・貯蔵中の変化」『聖霊女子短期大学紀要』第38巻、2010年、31–39頁、doi:10.24571/swjcb.38.0_31。
- ^ 井上友樹、村上拓彦、光田靖、宮島淳二、溝上展也、吉田茂二郎「ニホンジカによる人工林剥皮害と下層植生との関連性」『日本森林学会誌』第89巻第3号、日本森林学会、2007年、208–216頁、doi:10.4005/jjfs.89.208。
- ^ 『新常用和漢薬集[リンク切れ]』東京生薬協会。2020年3月28日閲覧。
- ^ 紙屋和宏、宇水泰三郎、島村雄三、橋本茂「キハダ優良品種の増殖と徳島県下におけるキハダ造林に関する調査研究」『徳島県林業総合技術センター研究報告』第38巻、2001年、6–13頁。
- ^ 巌佐 et al. 2013, p. 264, 「環状剥皮」.
- ^ Choi, C. (2008). “Tierra del Fuego: the beavers must die”. Nature 453: 968. doi:10.1038/453968a.
- ^ 浅野浩之、金澤好一、小野里光、鶴渕恒雄、綿貫邦男「ヒノキの巻き枯らし間伐実証試験」『研究報告』第14号、群馬県林業試験場、2009年、1-9頁、NAID 220000114953。
- ^ 小野里光、浅野浩之、金沢好一「ヒノキ巻枯らし間伐林における森林害虫の発生」『群馬林試研報』第14号、2009年、10–19頁、NAID 220000114954。
- ^ 崎尾均、川西基博、比嘉基紀、崎尾萌「特集「外来種と植生管理」 巻き枯らしによるハリエンジュの管理」『日本緑化工学会誌』第40巻第3号、2015年、446–450頁、doi:10.7211/jjsrt.40.446。
- ^ 水田泰徳「環状はく皮処理が主幹形モモの生育と果実品質に及ぼす影響」『兵庫県立農林水産技術総合センター研究報告〔農業編〕』第55号、2007年、5–9頁、NAID 40015454908。
- ^ 森口一志「キウイフルーツの生育期の環状はく皮による果実肥大効果、果実品質及び樹体への影響」『愛媛県立果樹試験場研究報告』第15巻、愛媛県立果樹試験場、2002年、55–65頁、NAID 220000084486。
- ^ 山根崇嘉、加藤淳子、柴山勝利「ブドウ‘安芸クイーン’の着色実態および環状はく皮と着果量の軽減による着色改善」『園芸学研究』第6巻第3号、2007年、441–447頁、doi:10.2503/hrj.6.441。
- ^ 長谷川耕二郎「カキ‘西村早生’の雄花と雌花の着生に及ぼす側枝の環状はく皮ならびに結縛の影響」『高知大学学術研究報告』第44巻、1995年、11–18頁、hdl:10126/833、NAID 120001107288。
- ^ 四方正義、星野正生、新城健、古沢寿治、クスウィアー・エンジャン「環状はく皮処理による亜熱帯系桑古条さし木の時期別露地実験」『日本蚕糸学雑誌』第55巻第2号、日本蚕糸学会、1986年、173–174頁、doi:10.11416/kontyushigen1930.55.173。
- ^ 池田虎三「各都道府県の林業・林産業と遺伝育種の関わり(24)石川県」『森林遺伝育種』第8巻第1号、森林遺伝育種学会、2019年、63–66頁、doi:10.32135/fgtb.8.1_63。
参考文献
[編集]- 巌佐庸、倉谷滋、斎藤成也 ほか 編『岩波 生物学辞典』(第5版)岩波書店、2013年。ISBN 978-4-00-080314-4。
- キャンベル、Urry、Cain、Wasserman、Minorsky、Reece『キャンベル生物学』池内昌彦、伊藤元己、箸本春樹、道上達男 監訳(原書11版)、丸善出版、2018年。ISBN 978-4621302767。
- 原襄『植物形態学』朝倉書店、1994年。ISBN 4-254-17086-6。
- ポーラ・ルダル 著、鈴木三男、田川裕美 訳『植物解剖学入門: 植物体の構造とその形成』八坂書房、1997年。ISBN 4-89694-696-0。
外部リンク
[編集]- 福原達人「1-3. 木部・篩部」『植物形態学』福岡教育大学。2020年3月7日閲覧。
- 日本光合成学会 編『光合成事典』。2020年3月28日閲覧。