ヨコエビ

| ヨコエビ亜目 |
|---|
 |
| 分類 |
ヨコエビ(横蝦、横海老)は、甲殻亜門・軟甲綱・端脚目(ヨコエビ目)に含まれ、ワレカラとクジラジラミを除くSenticaudata亜目に、Hyperiopsidea亜目,Amphilochidea亜目,Colomastigidea亜目の3亜目を加えたものを指す。かつてはヨコエビ亜目(Gammaridea)の総称であったが、2013年にヨコエビ亜目の一部のグループとワレカラ亜目(Caprelidea)を合せたSenticaudata亜目が設けられ(Lowry & Myers 2013)、その後ヨコエビ亜目がHyperiopsidea亜目,Amphilochidea亜目,Colomastigidea亜目の3亜目に分離され、現在に至る(Lowry & Myers 2017)。 以下、和名は 佐藤・伊藤 (1980),マグローヒル科学技術用語大辞典編集委員会 (1996),篠田 (2006),有山 (2022) に拠った。
端脚類の中でも特に種分化が進んだグループで、幅広い環境に多くの種が分布している。世界から9237種が知られており(Lowry & Myers 2017)、日本からは2021年現在で493種について確実な報告がある(有山 2022)。また、10,000m以深のチャレンジャー海淵においても生息が確認されている[1]。淡水にも、温帯や冷帯を中心に1800種以上が見つかっている。陸生のものはそれらに比べれば少ないが、それでも200種以上が海岸の草むらや森の落ち葉の下に生活している。
_front_view_-_Tyninghame_beach_-_East_Lothian,_Scotland_-_20_April_2013.jpg)
野外においてしばしば高い密度で生息するため、自然界では分解者として、また他の動物の餌として重要である。たとえば河口域において、ヨコエビ類が堆積した落ち葉を食べ分解すると同時に、魚類の餌となっている事例が知られている(櫻井 2004)。人間にとっての利用価値はほとんどないが、乾燥させたものがカメのカルシウム補給用の餌として販売されている。また、海水魚の飼育においてネズッポ類マンダリンやタツノオトシゴ・ヤッコなどが人工餌に餌付かない場合に生き餌として与えることがあるほか、食べかすの掃除等を目的として水槽内に繁殖させることもある。
形態[編集]

名称に「エビ」とあるが十脚目(エビ目)ではない。体長は種により異なり、数mmから十数cmに及ぶが、多くは数mm程度しかなく、1cmを超える種は限られている。ヨコエビ科(Gammaridae)など代表的な種において、体は左右に平たく、横から見ると半円形に似ている。エビ(十脚目)とヨコエビとの主な相違点として、以下の形態的特徴が挙げられる。
- エビ類は眼柄と背甲をもつが、いずれもヨコエビ類にはない
- エビ類の触角は全体が鞭状に伸長し、脚(歩脚)も全体が細長い
- エビ類のエラは高度に発達し羽毛状の構造をもつが、ヨコエビ類のそれは基本的に単純な袋状である
- エビ類では尾節の前半5節が遊泳脚をもった腹節となるが、ヨコエビ類は前半3節が腹節となる
- エビ類は腹部に卵を抱える構造をもつが、ヨコエビ類は胸部に抱卵する

2対の触角を具え、第1触角には3節,第2触角には5節の柄部があり、そこから数節~数十節に及ぶ主鞭が生じる。種により第1触角主鞭の根元に副鞭と呼ばれる短い鞭部を生じる。主鞭にヘラ状感覚毛(aesthetasc)やボタン状感覚器(calceolous),あるいはcallynophoreという器官をもつものが知られるが、種類によりこれを欠く(Bousfield 1994)。複眼は頭部の外骨格に埋没している。基本的に複眼は1対だが、中央部で癒合するもの(クチバシソコエビ科)、2対具えるもの(スガメソコエビ科)、個眼が分散傾向を示すもの(ナギサヨコエビ科の一部)、全く退化するもの(マミズヨコエビ科など暗居性種)が知られる。
第1,2胸脚(歩脚)は特に咬脚と呼ばれており、種によって様々な形状を見せる。左右非対称に発達するもの(メリタヨコエビ科の一部)や、全く消失するもの(マルハサミヨコエビ科の一部)も知られる。咬脚は多くが何かをつかむ機能を持っており、雄間闘争や捕食などに用いているとされる。ヨコエビ類のエラは各胸脚の底節板に生じるが、エラをもつ胸脚は種類により異なる。基本的に単純な袋型をしているが、種により表面に虫垂様の突起を生じる。また、胸節下側の中央部分に胸節鰓をもつグループも知られる。 メスが卵を抱える器官である覆卵葉も、底節板から生じる。
腹節には遊泳を行う腹肢を具える。腹肢には2つの副肢があり、羽毛状の剛毛を密生する。
尾節には、2つの副肢を具えた尾肢を1対ずつ具えるが、種により副肢は片方または両方が消失する。肛門は第3尾肢の付け根付近に位置し、その上には尾節板と呼ばれる器官がある。尾節板にはしばしば種の特徴が表れ、形態分類を行う上で重要である。
生態の多様性[編集]
生活様式[編集]
ヨコエビ類の多くは水生の底生生物だが、なかには遊泳するもの、さらには陸生のものもいる(大塚 & 駒井 2008)。
陸上生活をするハマトビムシ科の種は跳躍力に優れ、それぞれの和名も「~トビムシ」と名付けられている(トビムシ目(粘管目)の動物も「トビムシ」と呼ばれるが、同じ節足動物門ではあるものの亜門レベルで異なる別物である)。ハマトビムシ科のジャンプは脚ではなく、腹部を下に曲げてバネとする。高さは体の数十倍から100倍に達し、捕食者の眼をくらませるのに役立つ。
底質内に潜り込む習性をもつものや近底遊泳性の種には、「ソコエビ」との和名がつけられている。深海産は大型化する傾向があり、中には20数cmにも達する大型種もいる。

石の下などの隙間に棲むものは体を横に倒して生活することが多く、和名が「ヨコエビ」とつけられている。石をひっくり返すと腹部を激しく振って泳ぎだすが、深い水中ではふつうに体を立てて泳ぎ、再び石などの下に入り込む。これらの種は第5~7胸脚を三脚のように展開して平面上を歩き回ることができ、岩や海藻の表面も横向きのまま移動することがある。
被食[編集]
哺乳類[編集]
- バイカル湖には多数のバイカルアザラシが生息しているが、植物プランクトンに乏しく生産性の低い閉鎖水系でこれほど水生哺乳類が繁栄することは異例とされている。研究者がアザラシにカメラを取り付けて調査したところ、夜間に頻繁に潜水してヨコエビの一種Macrohectopus branickiiを1匹ずつ捕食し、その数は1日4300匹と見積もられた。バイカルアザラシの歯には特殊なギザギザ構造があり、小さなヨコエビを捕食するたびに歯の隙間から水を排出することで、効率よい捕食が可能になっていると推定される。魚類より栄養段階の低いヨコエビを捕食することが、栄養の乏しい環境でも水生哺乳類が繁栄できる要因の一つと考えられている[2]。
- 海底に潜むスガメソコエビ科がコククジラの主要な餌となっている(内田 1993)。
- 知床では潮間帯上部に生息するヨコエビ類がヒグマUrsus arctosの餌となっているとの報告がある[3]。
鳥類[編集]
- 渉禽類にとって、干潟の底質中に生息するヨコエビ類が重要な餌資源となっている。例えば、アカアシシギの主な餌としてドロクダムシ科の一種Corophium voltatorが挙げられる(Goss-Custard 1977)。
魚類[編集]
- 海水域および淡水域において、ヨコエビ類に高い選好性をもつ魚類が知られている。例えば、ヒラメ幼魚においてヨコエビ類(アゴナガヨコエビ科)に対して正の選択性が確認されている(片山 2007)。
- ビワマスの餌としてアナンデールヨコエビが圧倒的な割合を占めている(藤岡 2016)。
軟体動物[編集]
紐形動物[編集]
- 小笠原諸島より知られる外来種のヒモムシGeonemertes pelaensisが、ヨコエビ類(オカトビムシ類)やワラジムシ類などを捕食し、深刻な被害を与えていることが明らかになった(Shinobe et al. 2017)。これら陸棲甲殻類は、海洋島たる小笠原諸島においてその成立過程で定着しえなかったミミズに代わって土壌形成を担っていたため、外来ヒモムシが侵入した地域では植物由来遺物の分解が阻害されることによる物質循環の破綻が危惧されている。
その他[編集]
- ヨーロッパミドリガニCarcinus maenasに捕食されていることが明らかになっている(Baeta et al. 2006)。
- ニュージーランドに生息するヒトデの一種Stegnaster inflatusは、体の下側に入り込んだヨコエビを四方から腕を伸ばして口の周りまで追い詰めて捕食することが知られている(Grace 1974)。
.jpg)
寄生[編集]
- 鉤頭動物門の一種Polymorphus minutus (Zeder, 1800)は、淡水性のGammarus roeseli,Dikerogammarus villosus(いずれもヨコエビ科)を操作して水面近くまで遊泳させ、次の宿主に発見されやすくする(Médoc, Bollache & Beisel 2006)。鉤頭動物の寄生は海産ヨコエビにおいてもよく知られている。
- 貝形虫の仲間Acetabulastomaには、ヨコエビ類に寄生するものが報告されている(Hart 2013)。
- 微胞子虫の一種Fibrillanosema crangonycisは、Crangonyx pseudogracilis(マミズヨコエビ科)に寄生する(Johanna et al. 2004)。
- 線形動物門の一種Trophomera marionensisは、カイコウオオソコエビと近縁の深海性ヨコエビHirondellea dubiaに寄生する(Leduc & Wilson 2016)。
- グレガリナの仲間Rotundula gammari,Heliospora longissimaが、Gammarus fasciatus(ヨコエビ科)に寄生する(Grunberg 2017)。胞子虫の寄生は陸生ヨコエビにおいても報告されている。
- バイカル湖には、大型ヨコエビ類の覆卵葉内に寄生するヨコエビ類が知られている(Karaman 1976)。
行動様式および生理的特性[編集]

繁殖[編集]
ヨコエビ類はメスの産卵直後に体外受精を行い、卵および孵化直後の幼体はメスの覆卵葉内で一定期間保護される。トンガリドロクダムシにおいて三倍体と思われる雌雄の形質を具えた個体が発見されることがある(Chapman 2007)ものの、普通は雌雄異体である。他の小型甲殻類と同様、メスは脱皮直後の外骨格が柔らかいうちにしか産卵しないため、オスがメスを獲得する戦略は多様に発達している。典型的な様式は以下の通りである(Bellan-Santini 2015)(Beermann, Dick & Thiel 2015)。
探索[編集]
交尾前ガード[編集]
- 種に特有の方法によってメスを捕捉することで、受精可能なわずかな時間を逃さないようにする。
- ハマトビムシ上科,ヨコエビ上科,ハッジヨコエビ上科のように同じ向きで背面から抱え込むスタイルが一般的だが、マミズヨコエビ科やヨロイヨコエビ科は背面から交差するように捕捉する。
ハレム形成[編集]
- 雄間闘争によりオスが複数のメスを獲得し、支配下に置く。
- カマキリヨコエビ科では巣穴が密集したコロニー上で闘争が行われる。
- 砂粒や貝殻片でできた筒に入って移動する習性のあるヤドカリモドキ類においては、オスが自分の巣にメスの巣を合体させ、オスが複数のメスを持ち歩くようなかたちで独占する(ポータブル・ハレム)という、珍しい生態が知られている。
摂餌[編集]
ヨコエビ類の食性は植食,肉食,懸濁物食など多様である。ただし、特化した食性をもつグループは限定的であり、大部分のヨコエビ類は、水中に浸漬した動植物組織や有機砕屑物であれば何でも食べるいわば「掃除屋」の役割を果たしていると考えられている。以下に代表的な摂餌様式を示す。
植食者[編集]

捕食者[編集]
- クチバシソコエビ科は砂中に潜り、複眼が発達した頭部だけを水中へ出して獲物を探す。
- 日周鉛直運動を行う例が報告されているタテソコエビ科も捕食性を示す。
- ヨロイヨコエビ科は海底に潜み、獲物を待ち伏せて捕食する。

腐肉食者[編集]
- バイカル湖に生息するアカントガンマルス科のヨコエビ類は湖底を徘徊して魚類の死骸を漁ると考えられている。
- フトヒゲソコエビ上科は普段は底質中に潜んでおり、餌となる動物の死骸などが沈んでいると水中を遊泳して群がる。
- 人の水死体も、しばしばヨコエビによって損壊される[5]。
懸濁物食者[編集]
- 潜砂性ヨコエビの一部と造管性ヨコエビのほとんどが有機懸濁物を食べていると考えられる。
- ヨコエビ類は、底質の表面に堆積したり水の流れに乗って漂っている懸濁物を、触角や咬脚を用いて集める。また、腹肢を使って水流を起こし、棲管内部や底節板の隙間を通過させることによって、効率よく浮遊物を捕集しているとされている。この行動は摂餌のみならず、自身の呼吸や卵に新鮮な水を供給することにも役立っていると考えられている。
共生および寄生[編集]
- 刺胞動物,棘皮動物,外肛動物,海綿動物などの体表において、タテソコエビ科,テングヨコエビ科,ドロノミ科,ホテイヨコエビ科,ハッジヨコエビ上科などが発見されることが多い。また、内部への棲み込みも報告されており、尾索動物に対してホヤノカンノン属(エンマヨコエビ科)やマルハサミヨコエビ科が棲み込みを行うことが知られている。
- Hyachelia(モクズヨコエビ科)やPodocerus cheloniae(ドロノミ科)は、ウミガメの体表から得られており、生活の場としていることが知られている。
- ドロノミ科のジンベエドロノミPodocerus jinbeは、ジンベエザメの口腔内(鰓耙)に付着するという極めて特異的な習性をもつ(Tomikawa et al. 2019)。剛毛を密生した触角を使って懸濁物を捕集して摂食するタイプのヨコエビであり、魚の口の中という外敵のいない環境を巧みに利用している[6]。
- フトヒゲソコエビ上科サカテヨコエビ科のサカテヨコエビ属Trischizostomaは、深海魚に体表寄生することが知られている(Freire & Serejo 2004)。
- ニホンオカトビムシMorinoia japonicaはハランの送粉者となっていると考えられていたが(Kato 1995)、後の研究ではキノコバエ類が主にその役を担っているとの見解が示されている(Suetsugu & Sueyoshi 2017)。

アンフィポッド・シルク[編集]
- ヒゲナガヨコエビ科は、主に保育のため海藻をつづりあわせて巣を作る。ドロクダムシ科やカマキリヨコエビ科は、泥や砂の管を作り、その中に棲む。ホソツツムシ属CerapusやCyrtophiumは、緻密に作られた筒状の巣に入ったまま水中を泳いで移動する(Giles 1885)。ヤドカリモドキ類は巻貝の殻に小石や貝殻片をつなげてその中に棲み、巣をひきずるように移動する。シャクトリドロノミ科Dyopedos や Dulichiaはマスト状の構造物を形成し、子供を止まらせてしばらく同居する。これらを作成するために用いられる分泌物は第3,4胸脚にある分泌腺でつくられ、指節先端からセメント状ないし糸状になって排出されるため、アンフィポッド・シルクと呼ばれている。
発光[編集]
- Pseudorchestoidea gracilis(ハマトビムシ科)で生物発光が報告されている。ただし、独自の発光器官は持たず、ホタルエビ等と同様に発光バクテリアに由来すると推測されている(Bousfield & Klawe 1963)。なお、日本では栗本丹洲の記した『千虫譜』において、ヨコエビの一種と思われる生物が発光するとの記述が見られる[7]。
おもな種類[編集]

- オオオキソコエビ[8] Eurythenes gryllus
- 深海産の大型種で体長15cmほどにもなる。体色は赤で目は白色。カイコウオオソコエビと同様に体内に深海の水圧に対するように脂肪分を体内に多く貯え、この脂肪を海水よりも軽くして数千メートルの深海底よりもやや上方を泳いで、エサとなる生物の死体を捜す。
- ダイダラボッチ[8] Alicella gigantea
- ダイダラボッチ科 (Alicellidae)・ダイダラボッチ属(Alicella)。前種よりも更に大型となるヨコエビの中でも最大級の種類で、体長30cm以上にもなる巨大種であり、同じく数千メートルの深海底を体内脂肪分を利用して浮力調節し、深海に落ちてきた生物の遺体を食べる。体色は淡褐色。

- カイコウオオソコエビ Hirondellea gigas
- 世界でもっとも深い場所から見つかった動物の1つで、水深6000メートル以深の超深海底にのみ生息する(土田 2008)。
- ヒメハマトビムシ(種群)
- ハマトビムシ科(Talitridae)。体長は1cmほどで、体色は青灰色や赤灰色をしている。海岸の満潮線付近に多数生息し、ふだんは砂の中や石の下にもぐっている。流れ着いた海藻や動物の死骸などを餌としており、それらを持ち上げるとたくさんの個体がピョンピョンと飛び跳ねる。他のヨコエビ類と比較してかなり発達が弱いものの遊泳脚(腹肢)をもち、水中を泳ぐこともできる。従来ヒメハマトビムシと呼ばれてきた種には複数の種が含まれるが、識別するためには非常に微細な形質を検討する必要がある。
- オカトビムシ Morinoia humicola
- ハマトビムシ科(Talitridae)・モリノオカトビムシ属(Morinoia)。体長7-8mm。沼地や休耕田の周辺などに生息する陸生のヨコエビ。
- ニッポンヨコエビ Gammarus nipponensis
- ヨコエビ科(Gammaridae)・ヨコエビ属(Gammarus)。体長は1 cmほどで、体色は黄褐色をしている。水のきれいな川の、落ち葉や石の下に生息する。
- ディケロガマルス・ヴィロースス Dikerogammarus villosus (Killer shrimp)
- ヨコエビ科(Gammaridae)・Dikerogammarus属。カスピ海・黒海原産のヨコエビの仲間。
- アカントガンマルス ビクトリィ Acanthogammarus victorii
- Acanthogammaridae科。バイカル湖に固有のヨコエビで、淡水性ヨコエビの中では桁違いに大きく、背面に長く鋭いトゲをもっている。琵琶湖博物館ではロシアから譲り受けた個体を生体展示しており、世界で初めて幼体の養育に成功したことで話題となった[9]。
- キョウトメクラヨコエビ Pseudocrangonyx kyotonis
- メクラヨコエビ属(Pseudocrangonyx)。島根県,京都府などの洞窟や井戸などで発見されている(洞穴生物)。体長は数mmほどで、体は半透明の白色をしていて、名のとおり目が退化している。メクラヨコエビ属は四国や北海道などに複数種が生息しているほか、ユーラシア極東部にも分布する。
- フロリダマミズヨコエビ Crangonyx floridanus
- マミズヨコエビ属(Crangonyx)。1989年に古利根沼で確認されて以降、日本各地で発見が相次いでいる外来種。日本在来の淡水ヨコエビとは異なり、比較的水質の悪い場所でも生息できる。
分類[編集]
分類体系はLowry and Myers (2013, 2016, 2017, 2019, 2020)に基づき、その後の研究 - (Sidorov et al. 2018), (Ariyama & Hoshino 2020), (Alves, Lowry & Johnsson 2020)- で改訂された部分は個別に反映させた。
- Colomastigidea亜目
- Colomastigida下目
- Colomastigidira小目
- Colomastigoidea上科
- ツツヨコエビ科 Colomastigidae
- Colomastigoidea上科
- Pagetinidira小目
- Colomastigidira小目
- Colomastigida下目
- Hyperiopsidea亜目
- Amphilochidea亜目 - かつてのヨコエビ亜目Gammarideaに含まれていた分類群の大部分を引き継いでいる。
- Amphilochida下目
- Maxillipiidira小目
- Maxillipioidea上科
- アシマワシヨコエビ科 Maxillipiidae
- Maxillipioidea上科
- Oedicerotidira小目
- Oedicerotoidea上科
- Exoedicerotidae科
- クチバシソコエビ科 Oedicerotidae

ベルギー沿岸で採取されたクチバシソコエビ科の一種Monoculodes carinatus - ゲンコツヨコエビ科 Paracalliopiidae
- Oedicerotoidea上科
- Eusiridira小目
- テンロウ上科 Eusiroidea
- Bateidae科
- テンロウヨコエビ科 Eusiridae

アラスカで採取されたテンロウヨコエビ科の一種Eusirus holmi - Miramarassidae科
- Thurstonellidae科
- Liljeborgioidea上科
- トゲヨコエビ科 Liljeborgiidae
- Pseudamphilochidae科
- テンロウ上科 Eusiroidea
- Amphilochidira小目
- Amphilochoidea上科
- チビヨコエビ科 Amphilochidae

ベルギー沿岸で採取されたチビヨコエビ科の一種Apolochus neapolitanus - Bolttsiidae科
- Cressidae科
- ホテイヨコエビ科 Cyproideidae
- マキエホテイヨコエビ Metacyproidea makie
- Didymocheliidae科
- Nihotungidae科
- テングヨコエビ科 Pleustidae
- セバヨコエビ科 Sebidae
- ミジンヨコエビ科 Seborgiidae科
- タテソコエビ科 Stenothoidae

ベルギー沿岸で採取されたタテソコエビ科の一種Metopa rubrovittata
- チビヨコエビ科 Amphilochidae
- マルハサミヨコエビ上科 Leucothoidea
- マルハサミヨコエビ科 Leucothoidae
- Leucothoe eltoni

ベルギー沿岸で採取されたマルハサミヨコエビ科の一種Leucothoe incisa
- Leucothoe eltoni
- マルハサミヨコエビ科 Leucothoidae
- Iphimedioidea上科
- Acanthonotozomatidae科
- Acanthonotozomellidae科
- リュウコツヨコエビ科 Amathillopsidae
- Dikwidae科
- ヨロイヨコエビ科 Epimeriidae
- カッチュウヨコエビ科 Iphimediidae

ケープ半島で撮影されたカッチュウヨコエビ科の一種Iphimedia gibba - Lafystiidae科
- Laphystiopsidae科
- Ochlesidae科
- スベヨコエビ科 Odiidae
- ホムラスベヨコエビ Postodius igneus
- Sicafodiidae科
- Stilipedidae科
- Vicmusiidae科
- Amphilochoidea上科
- Maxillipiidira小目
- Lysianassida下目
- Synopiidira小目
- エンマヨコエビ上科 Dexaminoidea
- フタハナヨコエビ科 Atylidae

ベルギー沿岸で採取されたフタハナヨコエビ科の一種Atylus swammerdami - エンマヨコエビ科 Dexaminidae
- サカサヨコエビ科 Melphidippidae
- ミコヨコエビ科 Pardaliscidae
- フタハナヨコエビ科 Atylidae
- Synopioidea上科
- スガメソコエビ科 Ampeliscidae

スガメソコエビ科のクビナガスガメAmpelisca brevicornis - ヒヨリミヨコエビ科 Argissidae
- フクスケヨコエビ科 Synopiidae
- スガメソコエビ科 Ampeliscidae
- エンマヨコエビ上科 Dexaminoidea
- Haustoriidira小目
- Haustorioidea上科
- Cheidae科
- Condukiidae科
- ツノヒゲソコエビ科 Haustoriidae
- Ipanemidae科
- Otagiidae科
- ヒサシソコエビ科 Phoxocephalidae
- Phoxocephalopsidae科
- フタマタソコエビ科 Pontoporeiidae
- Priscillinidae科
- Sinurothoidae科
- Urohaustoriidae科
- マルソコエビ科 Urothoidae

ベルギー沿岸で採取されたマルソコエビ科の一種Urothoe brevicornis - Zobrachoidae科
- Haustorioidea上科
- Lysianassidira小目
- Alicelloidea上科
- フクレソコエビ上科 Stegocephaloidea
- フクレソコエビ科 Stegocephalidae

ベーリング海東部で採取されたフクレソコエビ科の一種
- フクレソコエビ科 Stegocephalidae
- フトヒゲソコエビ上科 Lysianassoidea
- Adeliellidae科
- Amaryllididae科
- Cebocaridae科
- ヒロハソコエビ科 Cyclocaridae
- ネコゼヨコエビ科 Cyphocarididae

ネコゼヨコエビ科の一種Cyphocaris richardi - オキソコエビ科 Eurytheneidae
- カイコウオオソコエビ科 Hirondelleidae
- カイコウオオソコエビ Hirondellea gigas

北海で採取されたフトヒゲソコエビ科の一種Lepidepecreum longicornis
- カイコウオオソコエビ Hirondellea gigas
- フトヒゲソコエビ科 Lysianassidae
- オオテソコエビ科 Opisidae
- クツミガキソコエビ科 Scopelocheiridae
- タカラソコエビ科 Tryphosidae
- ツノアゲソコエビ科 Uristidae
- Aristioidea上科
- Acidostomatidae科
- Ambasiidae科
- フカゾリソコエビ科 Aristiidae
- Conicostomatidae科
- Derjugianidae科
- チョッキリソコエビ科 Endevouridae
- Izinkalidae科
- Kergueleniidae科
- Lepidepecreellidae科
- チョウチンソコエビ科 Pakynidae
- Sophrosynidae科
- Thoriellidae科
- サカテヨコエビ科 Trischizostomatidae
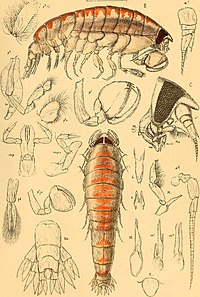
サカテヨコエビ属の一種Trischizostoma raschi - トウホクサカテヨコエビTrischizostoma tohokuense
- Wandinidae科
- Synopiidira小目
- Amphilochida下目
- Senticaudata亜目 - ワレカラ亜目を構成していたワレカラ類,クジラジラミ類が内包されている。以下の6下目からなる。
- Carangoliopsida下目
- Talitrida下目

バルト海に浮かぶボーンホルム島で撮影されたハマトビムシ科の一種Talitrus saltator - Talitridira小目
- Caspicoloidea上科
- Kurioidea上科
- Hyaloidea上科
- Ceinidae科
- Chiltoniidae科
- ナミノリソコエビ科 Dogielinotidae
- ネクイムシ科 Eophliantidae
- コンブノネクイムシ Ceinina japonica
- ヒアレラ科 Hyalellidae
- モクズヨコエビ科 Hyalidae

北海で採取されたモクズヨコエビ科の一種Apohyale prevostiiのオス - ヒョットコヨコエビ科 Najnidae
- ミノガサヨコエビ科 Phliantidae
- Plioplateidae科
- Temnophliantidae科
- ハマトビムシ上科 Talitroidea
- Talitridira小目
- Corophiida下目
- Corophiidira小目
- Aoroidea上科
- ユンボソコエビ科 Aoridae

ベルギー沿岸で採取されたユンボソコエビ科の一種Microdeutopus anomalusのオス - コタテヨコエビ科 Unciolidae
- ユンボソコエビ科 Aoridae
- Cheluroidea上科

キクイモドキChelura terebrans - キクイモドキ科 Cheluridae
- Chevalioidea上科
- ドロクダムシ上科 Corophioidea
- ヒゲナガヨコエビ科 Ampithoidae
- モズミヨコエビ Ampithoe valida
- ニッポンモバヨコエビ Ampithoe lacertosa

ニッポンモバヨコエビAmpithoe lacertosa
- ドロクダムシ科 Corophiidae

韓国で採取されたドロクダムシ科の一種Sinocorophium hangangenseのオス - トンガリドロクダムシMonocorophium insidiosum
- ヒゲナガヨコエビ科 Ampithoidae
- Aoroidea上科
- Caprelidira小目
- Aetiopedesoidea上科
- ワレカラ上科 Caprelloidea
- ワレカラ科 Caprellidae

ベルギー沿岸で採取されたワレカラの一種Pariambus typicus - ヨコエビワレカラ科 Caprogammaridae
- クジラジラミ科 Cyamidae
- シャクトリドロノミ科 Dulichiidae
- ドロノミ科 Podoceridae
- ワレカラ科 Caprellidae
- Isaeoidea上科
- イシクヨコエビ科 Isaeidae
- Microprotopoidea上科
- Neomegamphoidea上科
- Neomegamphopidae科
- キシドウヨコエビ科 Priscomilitariidae
- Photoidea上科
- カマキリヨコエビ科 Ischyroceridae
- カマカヨコエビ科 Kamakidae
- クダオソコエビ科 Photidae

クダオソコエビ科のフタトゲソコエビGammaropsis nitida
- ボウノボリヨコエビ上科 Protodulichioidea
- ボウノボリヨコエビ科 Protodulichiidae
- Rakirooidea上科
- Corophiidira小目
- Hadziida下目
- Hadziidira小目
- ハッジヨコエビ上科 Hadzioidea
- Crangoweckeliidae科
- センドウヨコエビ科 Eriopisidae
- Gammaroporeiidae科
- ハッジヨコエビ科 Hadziidae
- スンナリヨコエビ科 Maeridae
- メリタヨコエビ科 Melitidae
- Metacrangonyctidae科
- マルヨコエビ科 Nuuanuidae
- Calliopioidea上科
- ウラシマヨコエビ科 Calliopiidae
- Cheirocratidae科

ベルギー沿岸で採取されたウラシマヨコエビ上科Cheirocratidae科の一種Cheirocratus sundevallii - Hornelliidae科
- アゴナガヨコエビ科 Pontogeneiidae
- Magnovioidea上科
- ハッジヨコエビ上科 Hadzioidea
- Hadziidira小目
- Bogidiellida下目
- Bogidiellidira小目
- Bogidielloidea上科
- Artesiidae科
- カンゲキヨコエビ科 Bogidiellidae
- Salentinellidae科
- Bogidielloidea上科
- Bogidiellidira小目
- Gammarida下目
- Crangonyctidira小目
- Allocrangonyctoidea上科
- Crangonyctoidea上科
- Chillagoeidae科
- マミズヨコエビ科 Crangonyctidae

米国に生息するマミズヨコエビ科の一種Stygobromus kenki - Austroniphargidae科
- Giniphargidae科
- Kotumsaridae科
- シンヨコエビ科 Neoniphargidae
- Niphargidae科
- Paracrangonyctidae科
- Paramelitidae科
- Perthiidae科
- メクラヨコエビ科 Pseudocrangonyctidae
- Sandroidae科
- Sternophysingidae科
- Uronyctidae科
- Gammaridira小目
- ヨコエビ上科 Gammaroidea

バイカル湖の漁獲物の中にアカントガンマルス科とみられるヨコエビが混じっている - アカントガンマルス科 Acanthogammaridae
- アカントガンマルス・ビクトリィ Acanthogammarus victorii
- キタヨコエビ科 Anisogammaridae
- Baikalogammaridae科
- Bathyporeiidae科

ベルギー沿岸で採取されたヨコエビ上科Bathyporeiidae科の一種Bathyporeia guilliamsoniana - Behningiellidae科
- Falklandellidae科
- Gammaracanthidae科
- Gammarellidae科
- ヨコエビ科 Gammaridae

チェコに生息するヨコエビ科の一種Gammarus roeselii - ニッポンヨコエビ Gammarus nipponensis
- Iphigenellidae科
- コザヨコエビ科 Luciobliviidae
- Macrohectopidae科
- ナギサヨコエビ科 Mesogammaridae
- Micruropodidae科
- Ommatogammaridae科
- Pachyschesidae科
- Pallaseidae科
- Paraleptamphopidae科
- Phreatogammaridae科
- Pontogammaridae科
- Sensonatoridae科
- Typhlogammaridae科
- Zaramillidae科
- Zenkevitchiidae科
- アカントガンマルス科 Acanthogammaridae
- ヨコエビ上科 Gammaroidea
- Crangonyctidira小目
- Incertae sedis












_(16768822562).jpg)

.jpg)
.jpg)


.jpg)

.jpg)





人との関わり[編集]
環境科学[編集]

- アメリカ大陸に生息する淡水ヨコエビHyalella aztecaは、試験用生物として日本国内外において幅広く用いられている[10]。その他の種についても、有機化合物や重金属への応答について研究が行われている。
産業への影響[編集]
- 富山テレビ製作のドキュメンタリー番組『不可解な事実 ~黒部川ダム排砂問題~』では、2008年に富山県において網にかかったヒラメをヨコエビが食い荒らしているという映像が紹介され、出し平ダムからの排砂に含まれる有機物による富山湾の富栄養化が原因の一端とされる報道がなされた[11]。このヨコエビはフトヒゲソコエビ類の中のクツミガキソコエビ科に属するAroui onagawae (Takekawa & Ishimaru, 2000)であるが、大量発生のメカニズムについては明確になっていない部分が多い。
- 腐肉食性のヨコエビは魚の死骸をいちはやく分解する特性をもつ故に、えびかご漁業において生餌として仕掛けられているスケトウダラやニシンを完食し、ホッコクアカエビやトヤマエビの漁獲効率を低下させるとされている(飯田 et al. 2011)。
- 渓流に生息する淡水ヨコエビは、しばしばワサビを食害することが指摘されている。2009年には長野県安曇野で被害があり、オオエゾヨコエビJesogammarus jezoensisによるものと考えられている。また、島根県ではニッポンヨコエビGammarus nipponensisによる食害が指摘されているほか、静岡県でもヨコエビによる食害が報告されている。外来種であるフロリダマミズヨコエビCrangonyx floridanusもワサビに対する嗜好性が実験的に確かめられており(古屋 2011)、警戒の対象となっている[12]。
人体への被害[編集]
- 1965年、九州の志賀島沖にて、大きく損傷を受けた小型木造漁船が漂流しているのが発見され、衝突事故によるものと思われた。しかし、船内に人影はなく、直ちに周辺海域で乗員の捜索が行われた。翌日、近くの水深十数mから1体の遺体が発見されたが、着衣から露出した顔面や頸部を中心に激しく損傷しており、部分的に白骨化がみられるなど、死後かなりの日数が経過している様子であった。剖検を行ったところ、着衣の下には新鮮な組織が残存しており、行方不明となっていた乗員であると確認された。遺体の各所にはナイカイツノフトソコエビOrchomenella littoralis(フトヒゲソコエビ類)やウミホタルなどが付着・侵入していた。肺内部に多種の珪藻が見出されたことから死因は溺死であり、骨折痕がないことからスクリューでの創傷はなく、生物による摂食が行われて遺体が損壊したものと結論付けられた。また、後日加害船の航海日誌などから事故の詳細が調査され、これら小型生物が遺体頭部の白骨化に要した時間は、十数時間と見積もられた(永田, 福元 & 小嶋 1967)。その他、類似した事例として、タダノツノアゲソコエビAnonyx nugax,ヒメアナンデールヨコエビAnisogammarus fluvialis,トンガリキタヨコエビAnisogammarus pugettensisが関与したケースも報告されているが、白骨化には至っていない(小関 & 山内 1964)。ヨコエビやその他小型甲殻類による食害の初期には、粟粒大~小豆大の円形の食痕を生じ、それが進行するとあたかも切り取られたかのように部位の欠損がみられるようになるという。
- 2017年、オーストラリアの砂浜で冷たい海水に30分浸かっていた少年が、気づかない間に足首に無数の細かな傷を負い出血が止まらなくなるという事態に見舞われた[13]。医療機関では原因を突き止めることができなかったため、後日父親は現場で生肉を使用して容疑者となる生物を捕獲した。捕獲された生物は、専門家がフトヒゲソコエビ類と同定した[14]。このグループは腐肉食性を示すため、集団で餌を食べていた近くに長時間留まっていたことなどが被害の一因とも推測されているが、極めて珍しいケースと考えられている。なお、一部報道にてウミノミという名称が用いられているが、これはヨコエビ類ではなくクラゲノミ亜目を指す語であり、誤用である[15]。
デザイン[編集]
- ジャン・リュルサ(Jean Lurçat)のタペストリー「世界の歌 Le chant du Monde」に、ハマトビムシ科と思われるヨコエビ類が描かれている[16]。[要出典]
- アネット・チゾンとタラス・テイラーによる絵本バーバパパシリーズで、日本における2作目にあたる『バーバパパたびにでる』の中に、ヨコエビ類の挿絵が登場する(Tison & Taylor 1971)。[要出典]付属肢など各部の特徴は正確に描写されており、そこに書き込まれた物体の形状や配置は、鉤頭動物による寄生の様子を解説した図[17] に酷似している。[要出典]
- イギリス,フランス,ブルガリア,ペルー,オーストラリアなどでヨコエビ類があしらわれた切手が発行されている。特に、オーストラリアの切手にデザインされたヨコエビ類は後に「切手収集(philatelically)」にちなみAmaryllis philatelica Lowry & Stoddart, 2002と命名された。
その他[編集]

- 非常に長い学名をもつ動物として、Gammaracanthuskytodermogammarus loricatobaicalensis Dybowski, 1926やSiemienkiewicziechinogammarus siemenkiewitschii Dybowski, 1926が挙げられることがある。後者は動物命名法国際審議会への質問をきっかけに無効名とされた記録がある[18]ものの、いずれの学名も質問文中で原記載として指定されたベネディクト・ディボフスキィの論文[19]には登場しない。「非常に長い学名をもつヨコエビ」にまつわる豆知識は20世紀半ばから文献上で紹介されるようになり[20]、いくつかのバリエーションがあることから、そういったソースの孫引きにより生じた架空の生物名と考えるのが妥当である。
- エルトン・ジョンのファンであったJames Thomasは、マルハサミヨコエビ属の新種がもつ咬脚の特徴を、ピンボールの魔術師のミュージックビデオでエルトンが履いていたブーツに見立て、Leucothoe eltoni Thomas, 2015と命名した[21]。
- 2012年に放送されたTBSドラマATARUでは、海で発見された遺体の胃の中から甲殻類の成分が検出され、ヨコエビと一致したという描写がある。劇中では「スイーパー」というキーワードから、海の掃除屋と呼ばれていることが明かされ、"漁港で大量発生しているヨコエビ"の生体も登場する[22]。ドラマ内では、ヨコエビが死肉の分解に携わったことで検死段階での死亡推定時刻が1日程度長く見積もられたとされている。しかし実際は、ヨコエビの蚕食によって数日で白骨化が進み、死後数か月以上経過したような状態となることで死亡時期の判定が難しくなる、といった類のものである。
脚注[編集]
- ^ NHKスペシャル ディープ オーシャン超深海 地球最深(フルデプス)への挑戦 2017年8月27日(日)午後9時00分~9時49分放送
- ^ “バイカルアザラシのユニークな生態:わずか0.1グラムの小さな獲物を1匹ずつ食べていた”. 2020年11月17日閲覧。
- ^ 「知床ヒグマ調査 News Letter No.3」知床財団・知床博物館 平成26年(2014年)8月
- ^ えのすいトリーター日誌 メンダコチャレンジ2016 終章
- ^ #人との関わりを参照
- ^ “ジンベエザメの口内に新種のヨコエビ発見 広島大研究グループ”. 2019年10月28日閲覧。
- ^ 千蟲譜―国立国会図書館デジタルコレクション 三巻27頁 水蚤
- ^ a b “海洋生物(プランクトン) 分類データ LYSIANASSOIDEA”. 2016年1月10日閲覧。
- ^ 世界初 バイカルヨコエビの赤ちゃん誕生!!
- ^ 実験水生生物リスト(国立環境研究所) 2017年3月31日閲覧
- ^ 第19回FNSドキュメンタリー大賞ノミネート作品 2017年3月31日閲覧
- ^ “フロリダマミズヨコエビ 安曇野市役所”. 2018年8月19日閲覧。
- ^ The Washington Post「Flesh-eating sea bugs attacked an Australian teen’s legs: ‘There was no stopping the bleeding.’」 2017年8月11日アーカイブ
- ^ フトヒゲソコエビ類は、伝統的にフトヒゲソコエビ科とされていた一群で、現在はいくつもの科に分けられている。水死体から発見されたナイカイツノフトソコエビOrchomene naikaiensisやタダノツノアゲソコエビAnonyx nugax、そして刺し網漁業のヒラメを食い散らかしたとされるAuroi onagawaeも、同じフトヒゲソコエビ類に含まれる。
- ^ 端脚類#人との関わりを参照
- ^ Musées d'angers(全体画像)。Brodébrol(該当部分の拡大)。
- ^ Instructional Web Server The University of Western Ontario
- ^ “Opinion 105. Dybowski's (1926) Names of Crustacea Suppressed”, Opinions Rendered by the International Commission on Zoological Nomenclature: Opinions 105 to 114, Smithsonian Miscellaneous Collections, 73, (1929), pp. 1-3, hdl:10088/23619
- ^ Dybowski 1926 Bull. internat. Acad. Cracovie, (B) 1926:61.
- ^ Barbour, T. (1943), “The Sea and the Cave”, The Atlantic monthly: 99-103
- ^ New Species of Shrimp-Like Crustacean Named after Elton John SCI NEWS 2017年3月31日閲覧
- ^ "CASE3 身勝手な男心vs奇妙な女心". 日曜劇場 ATARU. 29 April 2012. TBS系。
{kind=link}
{kind=link}
参考文献[編集]
- Alves, J.; Lowry, J.; Johnsson, R. (2020), “A new superfamily and family of Hadziida (Amphipoda: Senticaudata), with a description of a new genus and new species from the Brazilian continental shelf”, Zootaxa 4184 (4), doi:10.11646/zootaxa.4779.4.8
- 有山啓之「ヨコエビとはどんな動物か?―形態・色彩・生態について―」『Cancer』第4779巻、121-126頁、2016年。doi:10.18988/cancer.25.0_121。
- 有山啓之『ヨコエビ ガイドブック』海文堂、2022年、160頁。ISBN 9784303800611。
- Ariyama, H.; Hoshino, O. (2020), “A new superfamily, family, genus and species of marine amphipod, Protodulichia scandens, from Japan (Crustacea: Amphipoda: Senticaudata: Corophiida)”, Journal of Natural History 53 (39-40): 2467-2477
- 朝倉彰 著「甲殻類とは」、朝倉彰 編『甲殻類学』東海大学出版会、2003年、27-28頁。ISBN 4486016114。
- Baeta, A.; Cabral, H; Marques, J.; Pardal, M. (2006), “Feeding ecology of the green crab, Carcinus maenas (L., 1758) in a temperate estuary, Portugal”, Crustaceana 79: 1181-1193
- Beermann, J.; Dick, J.; Thiel, M. (2015), “Social recognition in Amphipods: An overview”, in L. Aquiloni; E. Tricarico, Social Recognition in Invertebrates
- Bellan-Santini, D. (2015), “Order Amphipoda Latreille, 1816”, in C.V. Klein, Treatise on Zoology - Anatomy, Taxonomy, Biology. The Crustacea, Volume 5, Koninklijke Brill NV, pp. 93-248
- Bousfield, E.; Klawe, W. (1963), “Orchestoidea gracilis, a new beach hopper (Amphipoda: Talitridae) from lower California, Mexico, with remarks on its luminescence”, Bulletin, Southern California Academy of Sciences (Southern California Academy of Sciences) 62 (1): 6-8, doi:10.3160/0038-3872-62.1.1
- Bousfield, E.; Shih, C.-T. (1994), “The Phyletic Classification of Amphipod Crustaceans: Problems in Resolution”, Amphipacifica 1 (3): 76-134
- Chapman, John (2007), “Gammaridea”, in J.T. Carlton, The Light and Smith Manual Intertidal Invertibrates from Central California to Oregon. 4th edition, Universary California Press, pp. 545-618
- Freire, P.; Serejo, C. (2004), “The genus Trischizostoma (Crustacea: Amphipoda: Trischizostomidae) from the Southwest Atlantic, collected by the REVIZEE Program”, Zootaxa 645: 1-15
- 藤岡康弘「さけます情報 サケ科魚類のプロファイル-14 ビワマス」『SALMON情報』第10巻、49-52頁、2016年。
- 古屋, 洋一; 今津, 佳子; 久米, 一成; 金子, 亜由美 (2011), “静岡県における外来種(フロリダマミズヨコエビ)の生態調査” (PDF), 静岡県環境衛生科学研究所報告 54: 13-19
- Giles (1885), “Natural history notes form HM's Indian marine survey steamer" Investigator", commander Alfred Carpenter, RN commanding. No. 1. On the structure and habits of Cyrtophium calamicola, a new tubicolous amphipod from the Bay of Bengal”, Journal of the Asiatic Society of Bengal 54: 54-59
- Goss-Custard, J. (1977), “Predator responses and prey mortality in Redshank, Tringa totanus (L.), and a preferred prey, Corophium volutator (Pallas)”, Journal of Animal Ecology 46: 21-35, doi:10.2307/3944
- Grace, R. (1974), “Feeding behaviour of Stegnaster inflatus Hutton. (Class: Asteroidea, Family: Asterinidae).” (PDF), Tane 20: 162-165
- Grunberg, R.; Sukhdeo, M. (2017), “Temporal Community Structure in Two Gregarines (Rotundula gammari and Heliospora longissima) Co-Infecting the Amphipod Gammarus fasciatus”, Journal of Parasitology 103 (1): 6-13
- Hart, C. (2013), “A new species of parasitic ostracod of the genus Acetabulastoma (Paradoxostomatidae, Paradoxostominae) with a discussion of the copulatory appendage homologies”, Notulae Naturae 442: 1-11
- 飯田, 憲一 (2011) (PDF), えびかご漁業用人工蝟集餌料の開発 漁家経営安定を推進するえびかご漁業用ロングライフ人工蝟集餌料製造システムの開発(平成21~23年度)
- Ishimaru, Shin-ichi (1994), “A catalogue of gammaridean and ingolfiellidean Amphipoda recorded from the vicinity of Japan”, Report of the Sado Marine Biological Station (Niigata University) (24): 29-86
- Johanna, G.; Galbreath, S.; Smith, J.; Terry, R.; Becne, J.; Dunn, A. (2004), “Invasion success of Fibrillanosema crangonycis, n.sp., n.g.: a novel vertically transmitted microsporidian parasite from the invasive amphipod host Crangonyx pseudogracilis”, International Journal for Parasitology 34: 235-244
- Karaman, Godon (1976), “Contribution to the knowledge of the Amphipoda 80. Revision of the genus Pachyschesis Bazik. 1945 from Baikal Lake (Fam. Gammaridae)”, Poljoprivreda i Šumartvo 22 (4): 29-43
- 片山知史; 一色竜也; 張成年; 渡部諭史「相模湾におけるヒラメ種苗の摂食生態および摂食日周期性」『神水セ研報』第2巻、37-41頁、2007年。CRID 1050282813674903296。
- Kato, M. (1995), “The aspidistra and the amphipod”, Nature 377: 293, doi:10.1038/377293a0
- 小関, 恒雄; 山内, 峻呉 (1964), “水中死体の水生動物による死後損傷”, 日本法医学雑誌 18 (1): 12-20 (
 要購読契約)
要購読契約) - Leduc, D.; Wilson, J. (2016), “Benthimermithid nematode parasites of the amphipod Hirondellea dubia in the Kermadec Trench”, Parasitology Research 115: 1675-1682
- Lowry, James; Myers, Allan (2013), “A Phylogeny and Classification of the Senticaudata subord. nov.(Crustacea: Amphipoda) (Online edition)”, Zootaxa 3610 (1): 1-80, doi:10.11646, ISBN 978-1-77557-099-8
- Lowry, James; Myers, Allan (2016), “Zaramillidae, a new amphipod family from the subantarctic Kerguelen Islands (Amphipoda, Senticaudata, Gammaroidea, Zaramillidae fam. nov.)”, Zootaxa 4169 (2): 387-389, doi:10.11646/zootaxa.4169.2.11
- Lowry, James; Myers, Allan (2017), “A Phylogeny and Classification of the Amphipoda with the establishment of the new order Ingolfiellida (Crustacea: Peracarida)”, Zootaxa 4265 (1): 1-89, doi:10.11646/zootaxa.4265.1.1
- Lowry, James; Myers, Allan (2019), “New genera of Talitridae in the revised Superfamily Talitroidea Bulycheva 1957 (Crustacea, Amphipoda, Senticaudata)”, Zootaxa 4553 (1): 1-100
- Lowry, James; Myers, Allan (2020), “A phylogeny and classification of the Talitroidea (Amphipoda, Senticaudata) based on interpretation of morphological synapomorphies and homoplasies”, Zootaxa 4778 (2): 281-310
- マグローヒル科学技術用語大辞典編集委員会, ed. (1996), マグローヒル科学技術用語大辞典 (第3版), 日刊工業新聞社
- Médoc, V.; Bollache, L.; Beisel, J.-N. (2006), “Host manipulation of a freshwater crustacean (Gammarus roeseli) by an acanthocephalan parasite (Polymorphus minutus) in a biological invasion context”, International Journal for Parasitology 36 (13): 1351-1358
- 永田, 武明; 福元, 孝三郎; 小嶋, 亨 (1967), “フトヒゲソコエビ及びウミホタルによる水中死体損壊例”, 日本法医学雑誌 21 (5): 534-530, CRID 1520290884517148800 (要購読契約)
- 大塚攻; 駒井智幸 著「端脚目(Order Amphipoda)」、石川良輔編集 編『節足動物の多様性と系統』岩槻邦男・馬渡峻輔監修、裳華房〈バイオディバーシティ・シリーズ6〉、2008年、257-258頁。ISBN 9784785358297。
- 櫻井泉「森林が河口域の水産資源に及ぼす影響」(PDF)『北水試だより』第65巻、19-26頁、2004年。
- 佐藤隼夫; 伊藤猛夫『改訂 無脊椎動物採集・飼育・実験法』北隆館、東京、1980年。
- 篠田, 授樹 (2006), 東京都の湧水等に出現する地下水生生物の調査. 研究助成・一般研究, 28, 全国書誌番号:21265981
- Shinobe, S.; Uchida, S.; Mori, H.; Okochi, I.; Chiba, S. (2017), “Declining soil Crustacea in a World Heritage Site caused by land nemertean.”, Scientific Reports 7 (1), doi:10.1038/s41598-017-12653-4
- Sidorov, D.; Taylor, S.; Sharina, S.; Gontcharov, A. (2018), “Zenkevitchiidae fam. nov. (Crustacea: Gammaroidea), with description of new subterranean amphipods from extremely deep cave habitats”, Journal of Natural History 52 (23-24): 1509-153, doi:10.1080/00222933.2018.1482017
- Suetsugu, Kenji; Sueyoshi, M. (2017), “Subterranean flowers of Aspidistra elatior are mainly pollinated by not terrestrial amphipods but fungus gnats”, Ecology 98 (11), doi:10.1002/ecy.2021
- Tison, Annette; Taylor, Talus (1971) (French), Le voyage de Barbapapa, Paris: Les Livres di Dragon d'Or, ISBN 978-2-82120-129-3
- Tomikawa, K.; Yanagisawa, M.; Higashiji, T.; Yano, N.; Vader, W. (2019), “A New Species of Podocerus (Crustacea: Amphipoda: Podoceridae) Associated with the Whale Shark Rhincodon typus”, Species Diversity 24: 209-216, doi:10.12782/specdiv.24.20
- 土田真二 著「カイコウオオソコエビ」、藤倉克則・奥谷喬司・丸山正 編『潜水調査船が観た深海生物』東海大学出版会、2008年、253頁。ISBN 9784486017875。
- 内田安茂『わくわくウォッチング図鑑⑨ クジラ・イルカ』株式会社学習研究社、1993年。ISBN 4-05-200315-2。
| 典拠管理データベース: 国立図書館 |
|---|